And Can Reproduce Themselves. They Have Their Own Dna That Evolved From .
Cells are divided into two master classes, initially defined by whether they contain a nucleus. Prokaryotic cells (leaner) lack a nuclear envelope; eukaryotic cells have a nucleus in which the genetic material is separated from the cytoplasm. Prokaryotic cells are generally smaller and simpler than eukaryotic cells; in addition to the absenteeism of a nucleus, their genomes are less circuitous and they do not contain cytoplasmic organelles or a cytoskeleton (Table 1.1). In spite of these differences, the same basic molecular mechanisms govern the lives of both prokaryotes and eukaryotes, indicating that all present-day cells are descended from a single primordial antecedent. How did this kickoff cell develop? And how did the complexity and multifariousness exhibited past nowadays-twenty-four hour period cells evolve?
The Outset Cell
It appears that life first emerged at least iii.viii billion years agone, approximately 750 million years after Earth was formed (Figure 1.1). How life originated and how the first cell came into being are matters of speculation, since these events cannot be reproduced in the laboratory. Even so, several types of experiments provide important evidence begetting on some steps of the procedure.

Figure i.one
Time calibration of evolution. The scale indicates the gauge times at which some of the major events in the evolution of cells are thought to have occurred.
Information technology was first suggested in the 1920s that simple organic molecules could grade and spontaneously polymerize into macromolecules under the weather condition thought to exist in primitive Earth's atmosphere. At the time life arose, the atmosphere of Earth is idea to have contained little or no costless oxygen, instead consisting principally of CO2 and Northtwo in add-on to smaller amounts of gases such as H2, H2S, and CO. Such an temper provides reducing weather condition in which organic molecules, given a source of energy such equally sunlight or electric discharge, can form spontaneously. The spontaneous formation of organic molecules was first demonstrated experimentally in the 1950s, when Stanley Miller (so a graduate pupil) showed that the discharge of electric sparks into a mixture of H2, CH4, and NHthree, in the presence of water, led to the germination of a multifariousness of organic molecules, including several amino acids (Figure i.2). Although Miller's experiments did not precisely reproduce the conditions of primitive Earth, they clearly demonstrated the plausibility of the spontaneous synthesis of organic molecules, providing the basic materials from which the first living organisms arose.

Effigy 1.two
Spontaneous formation of organic molecules. Water vapor was refluxed through an atmosphere consisting of CH4, NHthree, and Hii, into which electric sparks were discharged. Assay of the reaction products revealed the formation of a diversity of organic molecules, (more than...)
The next step in development was the formation of macromolecules. The monomeric building blocks of macromolecules have been demonstrated to polymerize spontaneously under plausible prebiotic conditions. Heating dry mixtures of amino acids, for instance, results in their polymerization to form polypeptides. But the critical characteristic of the macromolecule from which life evolved must have been the ability to replicate itself. Simply a macromolecule capable of directing the synthesis of new copies of itself would have been capable of reproduction and further evolution.
Of the two major classes of advisory macromolecules in present-twenty-four hour period cells (nucleic acids and proteins), but the nucleic acids are capable of directing their own self-replication. Nucleic acids tin serve equally templates for their own synthesis as a result of specific base of operations pairing between complementary nucleotides (Figure 1.iii). A critical step in understanding molecular evolution was thus reached in the early on 1980s, when it was discovered in the laboratories of Sid Altman and Tom Cech that RNA is capable of catalyzing a number of chemical reactions, including the polymerization of nucleotides. RNA is thus uniquely able both to serve as a template for and to catalyze its own replication. Consequently, RNA is generally believed to have been the initial genetic system, and an early phase of chemical evolution is thought to have been based on self-replicating RNA molecules—a menstruum of evolution known equally the RNA world. Ordered interactions between RNA and amino acids so evolved into the present-day genetic lawmaking, and Dna somewhen replaced RNA as the genetic cloth.
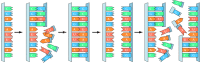
Figure ane.3
Self-replication of RNA. Complementary pairing between nucleotides (adenine [A] with uracil [U] and guanine [Grand] with cytosine [C]) allows one strand of RNA to serve every bit a template for the synthesis of a new strand with the complementary sequence.
The first cell is presumed to have arisen by the enclosure of cocky-replicating RNA in a membrane composed of phospholipids (Effigy one.4). Equally discussed in particular in the next chapter, phospholipids are the basic components of all present-day biological membranes, including the plasma membranes of both prokaryotic and eukaryotic cells. The key characteristic of the phospholipids that grade membranes is that they are amphipathic molecules, meaning that ane portion of the molecule is soluble in water and some other portion is not. Phospholipids take long, water-insoluble (hydrophobic) hydrocarbon bondage joined to water-soluble (hydrophilic) caput groups that incorporate phosphate. When placed in water, phospholipids spontaneously aggregate into a bilayer with their phosphate-containing head groups on the exterior in contact with water and their hydrocarbon tails in the interior in contact with each other. Such a phospholipid bilayer forms a stable barrier between two aqueous compartments—for example, separating the interior of the prison cell from its external surroundings.

Figure one.iv
Enclosure of self-replicating RNA in a phospholipid membrane. The kickoff cell is thought to take arisen past the enclosure of self-replicating RNA and associated molecules in a membrane composed of phospholipids. Each phospholipid molecule has two long hydrophobic (more...)
The enclosure of self-replicating RNA and associated molecules in a phospholipid membrane would thus have maintained them equally a unit of measurement, capable of self-reproduction and further evolution. RNA-directed protein synthesis may already have evolved by this time, in which case the showtime cell would have consisted of cocky-replicating RNA and its encoded proteins.
The Development of Metabolism
Because cells originated in a body of water of organic molecules, they were able to obtain food and energy directly from their surround. But such a situation is self-limiting, and then cells needed to evolve their own mechanisms for generating energy and synthesizing the molecules necessary for their replication. The generation and controlled utilization of metabolic energy is primal to all cell activities, and the principal pathways of free energy metabolism (discussed in detail in Affiliate 2) are highly conserved in present-twenty-four hours cells. All cells use adenosine 5′-triphosphate (ATP) as their source of metabolic free energy to drive the synthesis of cell constituents and acquit out other energy-requiring activities, such every bit movement (due east.grand., muscle contraction). The mechanisms used by cells for the generation of ATP are thought to have evolved in three stages, corresponding to the evolution of glycolysis, photosynthesis, and oxidative metabolism (Figure ane.5). The development of these metabolic pathways changed World'due south temper, thereby altering the course of further evolution.
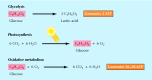
Figure one.5
Generation of metabolic free energy. Glycolysis is the anaerobic breakdown of glucose to lactic acrid. Photosynthesis utilizes free energy from sunlight to drive the synthesis of glucose from CO2 and HiiO, with the release of Otwo as a past-product. The O2 released past (more...)
In the initially anaerobic atmosphere of Earth, the kickoff energy-generating reactions presumably involved the breakdown of organic molecules in the absenteeism of oxygen. These reactions are likely to take been a form of present-day glycolysis—the anaerobic breakdown of glucose to lactic acid, with the net energy proceeds of two molecules of ATP. In addition to using ATP as their source of intracellular chemical energy, all present-day cells carry out glycolysis, consistent with the notion that these reactions arose very early in evolution.
Glycolysis provided a mechanism by which the energy in preformed organic molecules (due east.thou., glucose) could exist converted to ATP, which could then exist used every bit a source of energy to drive other metabolic reactions. The development of photosynthesis is generally thought to have been the next major evolutionary step, which allowed the cell to harness energy from sunlight and provided independence from the utilization of preformed organic molecules. The showtime photosynthetic bacteria, which evolved more than than 3 billion years ago, probably utilized HtwoS to catechumen COtwo to organic molecules—a pathway of photosynthesis still used past some bacteria. The utilise of HtwoO as a donor of electrons and hydrogen for the conversion of CO2 to organic compounds evolved later and had the important outcome of changing Earth's atmosphere. The use of H2O in photosynthetic reactions produces the past-product free O2; this mechanism is thought to take been responsible for making O2 abundant in Globe'due south temper.
The release of O2 as a event of photosynthesis inverse the environment in which cells evolved and is commonly thought to have led to the evolution of oxidative metabolism. Alternatively, oxidative metabolism may accept evolved before photosynthesis, with the increase in atmospheric O2 then providing a potent selective advantage for organisms capable of using Oii in energy-producing reactions. In either instance, Oii is a highly reactive molecule, and oxidative metabolism, utilizing this reactivity, has provided a machinery for generating energy from organic molecules that is much more than efficient than anaerobic glycolysis. For example, the consummate oxidative breakdown of glucose to CO2 and HiiO yields energy equivalent to that of 36 to 38 molecules of ATP, in contrast to the ii ATP molecules formed past anaerobic glycolysis. With few exceptions, present-day cells use oxidative reactions equally their main source of energy.
Present-Twenty-four hours Prokaryotes
Present-day prokaryotes, which include all the various types of bacteria, are divided into two groups—the archaebacteria and the eubacteria—which diverged early on in evolution. Some archaebacteria alive in extreme environments, which are unusual today but may have been prevalent in primitive Earth. For instance, thermoacidophiles live in hot sulfur springs with temperatures as high as lxxx°C and pH values as low as 2. The eubacteria include the common forms of present-day bacteria—a large grouping of organisms that live in a wide range of environments, including soil, h2o, and other organisms (east.g., man pathogens).
Virtually bacterial cells are spherical, rod-shaped, or spiral, with diameters of 1 to 10 μm. Their Dna contents range from about 0.6 1000000 to 5 1000000 base of operations pairs, an amount sufficient to encode about 5000 dissimilar proteins. The largest and well-nigh circuitous prokaryotes are the cyanobacteria, leaner in which photosynthesis evolved.
The construction of a typical prokaryotic cell is illustrated by Escherichia coli (E. coli), a common inhabitant of the human being intestinal tract (Figure 1.6). The cell is rod-shaped, virtually 1 μm in diameter and about 2 μm long. Like most other prokaryotes, East. coli is surrounded by a rigid cell wall composed of polysaccharides and peptides. Inside the cell wall is the plasma membrane, which is a bilayer of phospholipids and associated proteins. Whereas the prison cell wall is porous and readily penetrated by a diverseness of molecules, the plasma membrane provides the functional separation between the within of the cell and its external environment. The DNA of Due east. coli is a single circular molecule in the nucleoid, which, in contrast to the nucleus of eukaryotes, is not surrounded by a membrane separating it from the cytoplasm. The cytoplasm contains approximately 30,000 ribosomes (the sites of protein synthesis), which account for its granular appearance.

Figure 1.6
Electron micrograph of Due east. coli. The jail cell is surrounded by a cell wall, within which is the plasma membrane. DNA is located in the nucleoid. (Menge and Wurtz/Biozentrum, University of Basel/Science Photo Library/Photo Researchers, Inc.)
Eukaryotic Cells
Like prokaryotic cells, all eukaryotic cells are surrounded by plasma membranes and contain ribosomes. Still, eukaryotic cells are much more circuitous and incorporate a nucleus, a diverseness of cytoplasmic organelles, and a cytoskeleton (Effigy ane.seven). The largest and virtually prominent organelle of eukaryotic cells is the nucleus, with a bore of approximately v μm. The nucleus contains the genetic information of the prison cell, which in eukaryotes is organized as linear rather than circular Deoxyribonucleic acid molecules. The nucleus is the site of DNA replication and of RNA synthesis; the translation of RNA into proteins takes place on ribosomes in the cytoplasm.
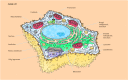
Figure 1.vii
Structures of animal and constitute cells. Both beast and establish cells are surrounded by a plasma membrane and incorporate a nucleus, a cytoskeleton, and many cytoplasmic organelles in mutual. Found cells are also surrounded by a cell wall and contain chloroplasts (more...)
In add-on to a nucleus, eukaryotic cells incorporate a variety of membrane-enclosed organelles within their cytoplasm. These organelles provide compartments in which unlike metabolic activities are localized. Eukaryotic cells are generally much larger than prokaryotic cells, ofttimes having a jail cell book at to the lowest degree a thousandfold greater. The compartmentalization provided by cytoplasmic organelles is what allows eukaryotic cells to function efficiently. Two of these organelles, mitochondria and chloroplasts, play disquisitional roles in free energy metabolism. Mitochondria, which are institute in nigh all eukaryotic cells, are the sites of oxidative metabolism and are thus responsible for generating nearly of the ATP derived from the breakdown of organic molecules. Chloroplasts are the sites of photosynthesis and are found only in the cells of plants and greenish algae. Lysosomes and peroxisomes too provide specialized metabolic compartments for the digestion of macromolecules and for various oxidative reactions, respectively. In improver, most plant cells contain large vacuoles that perform a variety of functions, including the digestion of macromolecules and the storage of both waste material products and nutrients.
Because of the size and complexity of eukaryotic cells, the send of proteins to their correct destinations within the cell is a formidable job. Two cytoplasmic organelles, the endoplasmic reticulum and the Golgi appliance, are specifically devoted to the sorting and transport of proteins destined for secretion, incorporation into the plasma membrane, and incorporation into lysosomes. The endoplasmic reticulum is an extensive network of intracellular membranes, extending from the nuclear membrane throughout the cytoplasm. Information technology functions not just in the processing and ship of proteins, simply also in the synthesis of lipids. From the endoplasmic reticulum, proteins are transported within minor membrane vesicles to the Golgi apparatus, where they are further processed and sorted for transport to their final destinations. In addition to this role in protein transport, the Golgi apparatus serves every bit a site of lipid synthesis and (in plant cells) as the site of synthesis of some of the polysaccharides that compose the cell wall.
Eukaryotic cells have another level of internal organisation: the cytoskeleton, a network of poly peptide filaments extending throughout the cytoplasm. The cytoskeleton provides the structural framework of the cell, determining cell shape and the general organization of the cytoplasm. In add-on, the cytoskeleton is responsible for the movements of unabridged cells (e.k., the wrinkle of musculus cells) and for the intracellular transport and positioning of organelles and other structures, including the movements of chromosomes during cell partition.
The eukaryotes adult at least 2.7 billion years ago, following some 1 to 1.5 billion years of prokaryotic evolution. Studies of their Dna sequences signal that the archaebacteria and eubacteria are as different from each other as either is from present-mean solar day eukaryotes. Therefore, a very early event in development appears to take been the divergence of three lines of descent from a mutual ancestor, giving rise to present-twenty-four hours archaebacteria, eubacteria, and eukaryotes. Interestingly, many archaebacterial genes are more than similar to those of eukaryotes than to those of eubacteria, indicating that the archaebacteria and eukaryotes share a common line of evolutionary descent and are more than closely related to each other than either is to the eubacteria (Figure ane.viii).
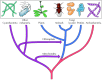
Effigy ane.8
Evolution of cells. Present-day cells evolved from a common prokaryotic ancestor along three lines of descent, giving rise to archaebacteria, eubacteria, and eukaryotes. Mitochondria and chloroplasts originated from the endosymbiotic association of aerobic (more...)
A critical step in the evolution of eukaryotic cells was the acquisition of membrane-enclosed subcellular organelles, allowing the development of the complication characteristic of these cells. The organelles are thought to have been acquired as a outcome of the association of prokaryotic cells with the ancestor of eukaryotes.
The hypothesis that eukaryotic cells evolved from a symbiotic association of prokaryotes—endosymbiosis—is particularly well supported past studies of mitochondria and chloroplasts, which are thought to accept evolved from bacteria living in large cells. Both mitochondria and chloroplasts are similar to bacteria in size, and like bacteria, they reproduce by dividing in 2. Well-nigh of import, both mitochondria and chloroplasts contain their own DNA, which encodes some of their components. The mitochondrial and chloroplast DNAs are replicated each fourth dimension the organelle divides, and the genes they encode are transcribed within the organelle and translated on organelle ribosomes. Mitochondria and chloroplasts thus contain their ain genetic systems, which are distinct from the nuclear genome of the jail cell. Furthermore, the ribosomes and ribosomal RNAs of these organelles are more closely related to those of bacteria than to those encoded by the nuclear genomes of eukaryotes.
An endosymbiotic origin for these organelles is now generally accustomed, with mitochondria thought to have evolved from aerobic bacteria and chloroplasts from photosynthetic bacteria, such as the blue-green alga. The acquisition of aerobic bacteria would have provided an anaerobic cell with the ability to carry out oxidative metabolism. The conquering of photosynthetic bacteria would have provided the nutritional independence afforded by the ability to perform photosynthesis. Thus, these endosymbiotic associations were highly advantageous to their partners and were selected for in the course of evolution. Through fourth dimension, nearly of the genes originally present in these leaner evidently became incorporated into the nuclear genome of the cell, and so just a few components of mitochondria and chloroplasts are still encoded past the organelle genomes.
The Evolution of Multicellular Organisms
Many eukaryotes are unicellular organisms that, like bacteria, consist of simply single cells capable of self-replication. The simplest eukaryotes are the yeasts. Yeasts are more than complex than leaner, but much smaller and simpler than the cells of animals or plants. For example, the ordinarily studied yeast Saccharomyces cerevisiae is about 6 μm in diameter and contains 12 million base pairs of DNA (Figure 1.9). Other unicellular eukaryotes, notwithstanding, are far more circuitous cells, some containing as much Dna as homo cells take (Table 1.2). They include organisms specialized to perform a variety of tasks, including photosynthesis, motility, and the capture and ingestion of other organisms as food. Amoeba proteus, for example, is a large, complex cell. Its volume is more 100,000 times that of Due east. coli, and its length can exceed ane mm when the cell is fully extended (Figure i.10). Amoebas are highly mobile organisms that use cytoplasmic extensions, called pseudopodia, to move and to engulf other organisms, including leaner and yeasts, every bit nutrient. Other unicellular eukaryotes (the green algae) contain chloroplasts and are able to carry out photosynthesis.

Figure 1.9
Scanning electron micrograph of Saccharomyces cerevisiae. Artificial color has been added to the micrograph. (Andrew Syed/Science Photo Library/ Photograph Researchers, Inc.)

Figure i.10
Light micrograph of Amoeba proteus. (Thou. I. Walker/Photo Researchers, Inc.)
Multicellular organisms evolved from unicellular eukaryotes at to the lowest degree 1.7 billion years ago. Some unicellular eukaryotes form multicellular aggregates that announced to correspond an evolutionary transition from unmarried cells to multicellular organisms. For instance, the cells of many algae (due east.g., the green alga Volvox) associate with each other to class multicellular colonies (Figure 1.11), which are idea to have been the evolutionary precursors of present-day plants. Increasing cell specialization then led to the transition from colonial aggregates to truly multicellular organisms. Continuing cell specialization and partitioning of labor among the cells of an organism accept led to the complexity and diversity observed in the many types of cells that make upwardly nowadays-day plants and animals, including human beings.

Figure i.11
Colonial green algae. Private cells of Volvox course colonies consisting of hollow balls in which hundreds or thousands of cells are embedded in a gelatinous matrix. (Cabisco/Visuals Unlimited.)
Plants are equanimous of fewer cell types than are animals, but each different kind of plant jail cell is specialized to perform specific tasks required by the organism as a whole (Figure 1.12). The cells of plants are organized into three master tissue systems: ground tissue, dermal tissue, and vascular tissue. The ground tissue contains parenchyma cells, which carry out most of the metabolic reactions of the establish, including photosynthesis. Ground tissue as well contains 2 specialized cell types (collenchyma cells and sclerenchyma cells) that are characterized by thick prison cell walls and provide structural support to the plant. Dermal tissue covers the surface of the plant and is equanimous of epidermal cells, which course a protective coat and allow the absorption of nutrients. Finally, several types of elongated cells class the vascular system (the xylem and phloem), which is responsible for the transport of water and nutrients throughout the found.

Figure 1.12
Lite micrographs of representative plant cells. (A) Parenchyma cells, which are responsible for photosynthesis and other metabolic reactions. (B) Collenchyma cells, which are specialized for back up and have thickened jail cell walls. (C) Epidermal cells (more...)
The cells found in animals are considerably more diverse than those of plants. The human body, for example, is composed of more 200 different kinds of cells, which are generally considered to be components of five main types of tissues: epithelial tissue, connective tissue, blood, nervous tissue, and muscle (Figure 1.13). Epithelial cells form sheets that encompass the surface of the body and line the internal organs. In that location are many different types of epithelial cells, each specialized for a specific function, including protection (the skin), assimilation (e.grand., the cells lining the pocket-sized intestine), and secretion (east.grand., cells of the salivary gland). Connective tissues include bone, cartilage, and adipose tissue, each of which is formed by unlike types of cells (osteoblasts, chondrocytes, and adipocytes, respectively). The loose connective tissue that underlies epithelial layers and fills the spaces between organs and tissues in the body is formed by another jail cell type, the fibroblast. Blood contains several different types of cells, which part in oxygen transport (crimson blood cells, or erythrocytes), inflammatory reactions (granulocytes, monocytes, and macrophages), and the immune response (lymphocytes). Nervous tissue is composed of nerve cells, or neurons, which are highly specialized to transmit signals throughout the body. Various types of sensory cells, such as cells of the eye and ear, are further specialized to receive external signals from the environment. Finally, several dissimilar types of musculus cells are responsible for the production of force and motility.

Figure 1.13
Light micrographs of representative animal cells. (A) Epithelial cells of the mouth (a thick, multilayered sheet), bile duct, and intestine. (B) Fibroblasts are connective tissue cells characterized by their elongated spindle shape. (C) Erythrocytes, (more...)
The evolution of animals conspicuously involved the development of considerable diverseness and specialization at the cellular level. Understanding the mechanisms that control the growth and differentiation of such a complex assortment of specialized cells, starting from a unmarried fertilized egg, is i of the major challenges facing contemporary prison cell and molecular biology.
Source: https://www.ncbi.nlm.nih.gov/books/NBK9841/
0 Response to "And Can Reproduce Themselves. They Have Their Own Dna That Evolved From ."
ارسال یک نظر